Ergebnisse::Vegetationseinheiten::
VegetationDie Artenliste der Farn- und Bl¸tenpflanzen (Anhang1) enth‰lt alle Taxa der vegetationskundlichen Aufnahmen aus den Jahren 1997 und 1998 auf den Untersuchungsfl‰chen bzw. den Transekten. Insgesamt konnten 119 Pflanzenarten nachgewiesen werden, wovon f¸nf Arten in Rheinland-Pfalz (Hand 1994) und zehn Arten in Nordrhein-Westfalen (Wolff-Straub et al. 1988) in der Roten Liste stehen (Tab. 8). Von diesen zehn Arten werden vier in der Eifel als gef‰hrdet eingestuft. 42 Arten wurden ausschlieþlich in Deutschland nachgewiesen, davon sind jedoch nur drei Arten (ca. 7 %) einem Gef‰hrdungsgrad sowohl in Nordrhein-Westfalen als auch in Rheinland-Pfalz zuzuordnen. Anders sieht es auf der belgischen Seite aus. Hier wurden zwar nur 21 Arten nachgewiesen, die ausschlieþlich in Belgien gefunden wurden, davon sind aber neun Arten (ca. 42 %) in ihrem Bestand gef‰hrdet. Wie es um den Gef‰hrdungsstatus der nachgewiesenen Pflanzenarten in Belgien, bzw. im belgischen Teil der Eifel steht, konnte mangels einer Belgischen Roten Liste nicht festgestellt werden. Tab. 8 : Gef‰hrdete Pflanzenarten und deren Vorkommen im Untersuchungsgebiet.
VegetationseinheitenEs wurden 13 unterschiedliche Vegetationseinheiten ermittelt. Die Pflanzendecke der Untersuchungsfl‰chen in den Bacht‰lern bildet oft ein kleinfl‰chiges Mosaik von nebeneinander liegenden Gesellschaften unterschiedlicher pflanzensoziologischer Gruppen. Sie werden beeinfluþt durch das M‰andrieren des Baches, durch n‰hrstoffreiche Hangw‰sser und durch das Auftreten von Quellen oder Quellmooren im Bachtalbereich. Die Vegetation der Feuchtheiden und Heidemoore ist ¸ber groþe Fl‰chen gleichfˆrmiger und weniger artenreich. Aber auch hier kommt es, oft durch anthropogene Einfl¸sse, zu kleinr‰umigen ƒnderungen der Vegetation.
Synsystematische Ðbersicht der ermittelten Vegetationseinheiten (nach Oberdorfer 1994)O. = Ordnung V. = Verband A. = Assoziation
FalterfaunaArtenspektrumIn den Jahren 1998 und 1999 konnten bei 188 Fl‰chenbegehungen 946 Falterbeobachtungen von insgesamt 32 tagaktiven Schmetterlingsarten gemacht werden (Tab. 9). Tab. 9: Anzahl
der Flächenbegehungen, Artenzahl und Anzahl der Beobachtungen
in den Jahren 1998 und 1999
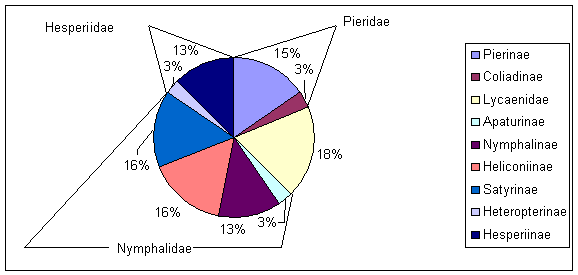
Aus dieser Tabelle wird ersichtlich, daß im Jahre 1999 wesentlich weniger Falterarten gefunden wurden. Dies liegt zum einen daran, daß die Begehungen in diesem Jahr nur von Mai bis Juli stattfanden, da sie sich auf die Hauptflugzeit von Lycaena helle und Boloria eunomia konzentrierten. Viele Arten, deren Hauptflugzeit später im Jahreslauf liegt, konn-ten so nicht erfaßt werden. Zum anderen schwanken die Falterpopulationen von Jahr zu Jahr sehr stark und bei einigen Falterarten ist die Nachweisbarkeit sehr stark zufallsbedingt (POLLARD & YATES 1993). Weder im Jahre 1998 noch im Jahre 1999 wurden Falter aus der Familie der Zygaenidae nachgewiesen. Die Nymphalidae mit den Unterfamilien Apaturinae, Nymphalinae, Heliconiinae und Satyrinae stellten mit 15 Vertretern die artenreichste Familie dar (Abb. 10). Die Lycaenidae und die Pieridae sind mit je 6 Arten vertreten, die arten‰rmste Gruppe sind die Hesperiidae mit 5 Vertretern.
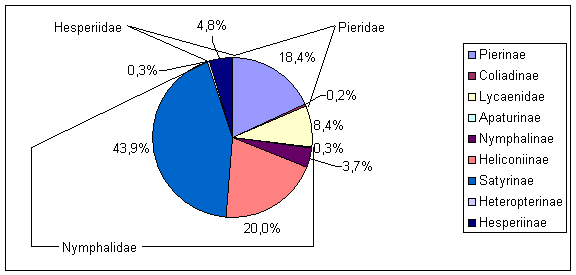
Bei der Verteilung der Individuen der einzelnen Arten auf die Familien bzw. Unterfamilien ergibt sich ein anderes Bild (Abb. 11).
Die Nymphalidae stellen hier mit 438 Individuen über zwei Drittel der gezählten Falter. Dabei machen die Satyrinae mit 283 Individuen die größte Gruppe aus. Der Anteil der Co-liadinae, der Apaturinae und der Heteroptinae ist mit unter 1 % verschwindend gering. Von den erfaßten 32 Arten waren 18 Arten mit weniger als zehn Individuen vertreten, bei sieben Arten handelt es sich um Einzelnachweise (Tab. 10).
Tab. 10: Übersicht der erfaßten Tagfalter der Jahre 1998 und 1999
Gefährdung der erfassten ArtenVon den 32 erfaþten Arten sind 15 (rund 47 %) einer Gef‰hrdungsstufe zuzuordnen. Davon sind f¸nf Falterarten deutschlandweit (Pretscher et al. 1998), elf Arten in Rheinland-Pfalz (Bl‰sius et al. 1992) und zwˆlf Arten in Nordrhein-Westfalen gef‰hrdet (Kinkler et al. 1988). F¸r das Gebiet Eifel-Siebengebirge werden neun Arten als gef‰hrdet eingestuft (Tab. 11). Davon ist Boloria aquilonaris (Hochmoor-Perlmutterfalter) akut vom Aussterben bedroht. Auch in Belgien wird diese Art als in ihrem Bestand gef‰hrdet eingestuft (Baguette & Goffart 1991, Baguette et al. 1992). Von Boloria aquilonaris wurden sechs Individuen auf zwei Untersuchungsfl‰chen nachgewiesen. Bei dem Fund auf einer Fl‰che handelt es sich um einen Einzelnachweis. Nach Wagener [1] wurden im Jahre 1998 aber noch mehrere Exemplare dort beobachtet. Bei den Arten Callophrys rubi, Celastrina argiolus, Polyommatus bellargus, Argynnis paphia und Melanargia galathea handelt es sich um Einzelnachweise. Apatura iris wurde mit lediglich zwei Exemplaren auf einer Untersuchungsfl‰che (Groþweberbach 1, Belgien) gefunden. Lycaena helle gilt sowohl deutschlandweit als auch in Nordrhein-Westfalen und Rheinland-Pfalz als akut vom Aussterben bedroht. Im Untersuchungsgebiet wurden insgesamt 93 Individuen dieser Art nachgewiesen. Es scheint also noch eine recht starke Population im Bereich der Hocheifel zu bestehen. Darauf weist auch der Gef‰hrdungsstatus f¸r den Bereich Eifel-Siebengebirge hin, der bei 2 (gef‰hrdet) liegt. Lycaena helle wurde sowohl auf Fl‰chen in Nordrhein-Westfalen (sechs von neun) und Rheinland-Pfalz (vier von vier) als auch auf belgischen Fl‰chen (f¸nf von 14) nachgewiesen. Boloria eunomia wird in allen Gebieten als gef‰hrdet eingestuft. Von dieser Art wurden insgesamt 108 Individuen auf zwˆlf Fl‰chen (sieben in Belgien, eine in Nordrhein-Westfalen und vier in Rheinland-Pfalz) gefunden. Sowohl Lycaena helle als auch Boloria eunomia gelten in Belgien als gef‰hrdet (Baguette & Goffart, 1991, Baguette et al. 1992). Brenthis ino und Boloria selene, die deutschlandweit lediglich auf der Vorwarnliste stehen wurden in wesentlich geringeren Individuenzahlen (41 bzw. 51) gefunden. Auffallend ist die geringe Individuenzahl von Lycaena hippothoe (8), dessen typische Lebensr‰ume Hangmoore und nasse Wiesen sind (Tolman & Lewington 1998). Tab. 11: Gefährdete Arten im Untersuchungsgebiet (1. & 2. nach KINKLER et al. 1988; 3. nach BLÄSIUS et al. 1992; 4. Nach PRETSCHER et al. 1998); türkis unterlegt: nur in Belgien nachgewiesen; gelb unterlegt: nur in NRW nachgewiesen; grün unterlegt: nur in RLP nachgewiesen
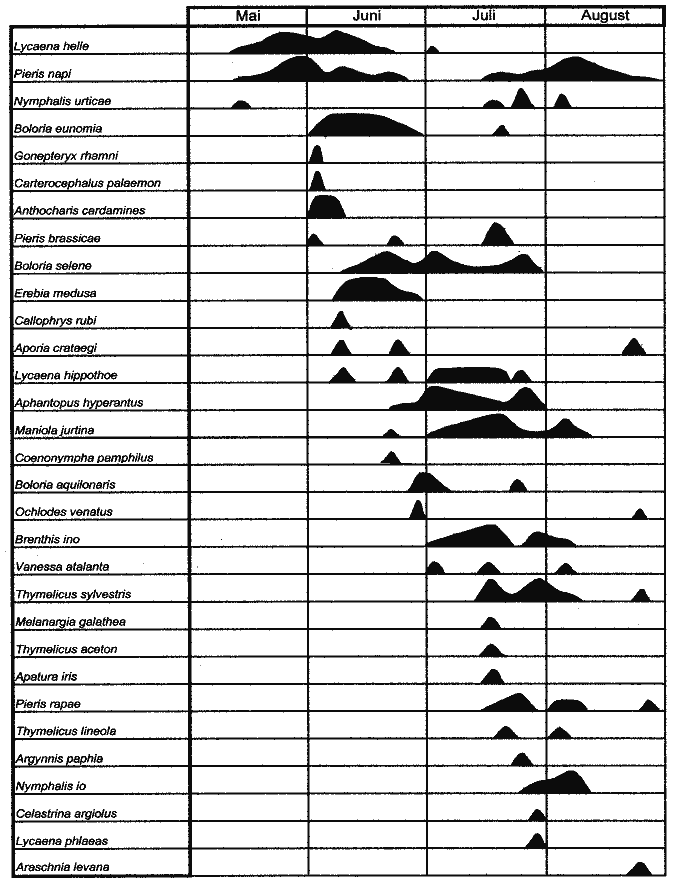
Ph‰nologieIn Abb.Ý12 sind die Nachweise der erfaþten Arten im Untersuchungszeitraum 1998 dargestellt.
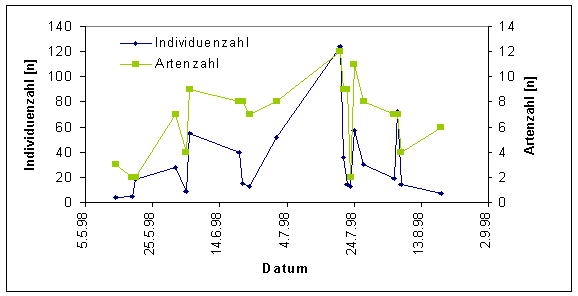
Die längste Flugzeit mit 95 Tagen wurde bei Pieris napi beobachtet. Diese Art flog während des gesamten Untersuchungszeitraumes. Bei Lycaena helle begann die Flugzeit am 14.5.98 und endete mit der letzten Beobachtung am 1.7.98. Insgesamt flog die Art also etwas mehr als sechs Wochen. Boloria eunomia wurde am 1.6.98 erstmals gefunden, der letzte Nachweis gelang am 21.7.98. Damit betrug die Gesamtflugzeit knapp acht Wochen. Das Auftreten von Lycaena helle und Boloria eunomia überschneidet sich also zeitlich in einem Rahmen von 30 Tagen. Bei beiden Arten war die Individuendichte Anfang Juni am höchsten (L. helle - 15 Falter; B. eunomia - 23 Falter). Sowohl die höchste Individuendichte als auch die größte Artenanzahl wurden im Jahre 1998 Ende Juli erreicht. Hier wurden 120 Individuen aus zwölf Arten gezählt (Abb. 13). Es handelt sich hier um die Hauptflugzeit der beiden individuenreichsten Arten Maniola jurtina und Aphantopus hyperantus. Ausgesprochene Frühjahrsarten sind hier wegen des relativ späten Beginns der Untersuchung am 14.5.98 unterrepräsentiert.
Präsenz, DominanzDie Falter Aphantopus hyperantus, Pieris napi, Maniola jurtina und Brenthis ino treten im Untersuchungsgebiet als dominante Arten auf (Abb. 14).
Abb. 14: Dominanz und Präsenz der erfaßten Arten im Untersuchungsgebiet (dunkelgrün: auf Feuchtstandorte spezialisierte Arten) 1998 Jeweils vier Arten sind dominant bzw. subdominant und machen rund 78 % der Beobach-tungen aus. Diesen acht Hauptarten stehen die restlichen als Begleitarten gegenüber. Bei Pieris napi handelt es sich um einen azönen Wanderfalter. Aphantopus hyperantus, Maniola jurtina und Thymelicus sylvestris gehören nach WEIDNER (1992) zu den eurytopen Begleitern ohne ausgesprochene Habitatpräferenzen. Aber auch diese Arten (insbesondere Maniola jurtina) sind in den letzten Jahren seltener geworden (WEIDEMANN 1995) und durch die zunehmende Intensivierung der Flächennutzung bedroht. Sie indizieren somit eine höhere Biotopqualität, hinsichtlich der Artenvielfalt. Zu den typischen Feuchtwiesenvertretern unter den Tagfaltern gehören Lycaena helle und Boloria eunomia. Diese Arten gelten als extrem stenotop und monophag; ihre Raupen sind auf Polygonum bistorta (Schlangenknöterich) als ausschließliche Futterpflanze spezialisiert. Ebenfalls auf feuchten Flächen sind Boloria selene und Brenthis ino anzutreffen. Die Raupen von Boloria selene fressen an verschiedenen Veilchen-Arten (Viola canina (Hunds-Veilchen), V. palustris (Sumpf-V.), V. riviniana (Hain-V.), V. hirta (Rauhhaar-V.). Brenthis ino-Raupen sind auf Filipendula ulmaria (Mädesüß) als Futterpflanze angewiesen (WEIDNER 1992). Nach TOLMAN & LEWINGTON (1998) und WEIDEMANN (1995) fressen sie auch an Sanguisorba officinalis (Großer Wiesenknopf) und anderen Pflanzenarten, die aber im Untersuchungsgebiet nicht gefunden wurden und damit hier keine Rolle spielen. Brenthis ino wird von WEIDNER aufgrund der Abhängigkeit von Filipendula ulmaria als zönobiont eingestuft, die Nutzung weiterer Pflanzenarten als Raupenfutter könnte diese Einstufung als zu eng erscheinen lassen. Ebenfalls in die Kategorie der Zönobionten fallen die oben genannten Arten Boloria eunomia und Lycaena helle. Die Falter Anthocharis cardamines und Lycaena hippothoe werden ebenso wie Boloria selene von WEIDNER (1992) als zönophil eingestuft, da sie auch in anderen Gesellschaften als den hier ermittelten vorkommen. Die geringe Präsenz der Arten Brenthis ino und Boloria eunomia im Verhältnis zu ihrem relativ hohen Dominanzindex deuten auf eine starke Bindung an bestimmte Habitatstrukturen hin. Bei Brenthis ino ist dieses Verhältnis am stärksten ausgeprägt (40 Individuen in zehn Rastern bzw. sieben Transekten), bei Boloria eunomia (55 Individuen in 13 Rastern bzw. elf Untersuchungsflächen). Lycaena helle hingegen wurde in weit mehr Rastern (21) und Untersuchungsflächen (15) angetroffen. Bei Boloria aquilonaris fällt eine Bewertung aufgrund der geringen Individuenzahl (sechs) wie bei den anderen Begleitarten schwer. Da die Art jedoch nur auf zwei Flächen nachgewiesen wurde, kann man von einer stark ausgeprägten Bindung der Art an spezielle Habitate ausgehen, zumal die ausschließliche Raupenfutterpflanze Oxycoccus palustris (Gemeine Moosbeere) ist. Der Bestand dieser Art ist in den letzten Jahren extrem zurückgegangen (PRETSCHER et al. 1998) und eine gezielte Untersuchung der tatsächlichen und der potentiellen Lebensräume noch nicht bekannter Vorkommen wird dringend empfohlen (MEINECKE 1985).
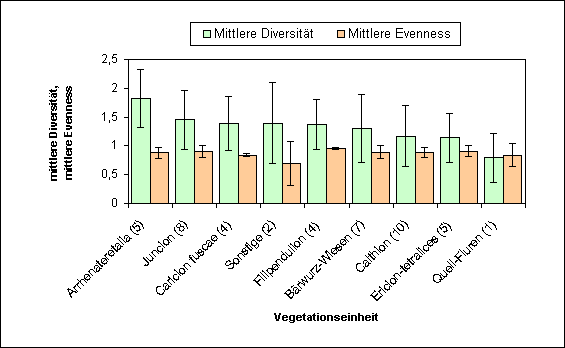
Verh‰ltnis von Vegetation und FalterfaunaDiversit‰t, EvennessDie Diversit‰t der Falterfauna auf den unterschiedlichen Vegetationseinheiten reicht von HS = 1,82 auf den Arrhenatheretalia-Gesellschaften bis HS = 0,79 auf den Quell-Fluren. Bei den anderen Fl‰chen sind die Unterschiede nicht so stark. Auch die Fl‰chen, die aufgrund ihrer Nutzung keiner Vegetationseinheit zuzuordnen sind (Sonstige), liegen im mittleren Bereich.
Diese Fl‰chen fallen bei der Evenness jedoch deutlich zur¸ck (ES = 0,7). Der hˆchste Wert wird hier von der Falterfauna der Filipendulion-Gesellschaften (ES = 0,95) erreicht. Bei den ¸brigen Fl‰chen ist die Evenness recht ausgeglichen (von ES = 0,84 bis ES = 0,9). Auffallend ist, daþ die Quell-Fluren trotz niedriger Diversit‰t eine hohe Evenness erreichen (ES = 0,84). Allgemein l‰þt sich sagen, daþ die Fl‰chen mit hoher Evenness eine relativ gleichm‰þige Verteilung der Individuen auf die Arten aufweisen. Das heiþt, daþ keine ausgesprochen dominanten aber auch nicht viele rezedenten Arten auf diesen Fl‰chen vorhanden sind. Die Kurve der Dominanzstruktur verl‰uft hier sehr flach, es herrscht eine st‰rkere Ausgeglichenheit zwischen den Individuenzahlen der Arten. Eine hohe Diversit‰t weist zwar auf eine groþe Artenanzahl hin, l‰þt aber die Verteilung der Individuen auþer acht. Ein Zusammenhang mit der Anzahl der Rote Liste-Arten hinsichtlich der Vegetationseinheiten wurde nicht festgestellt. Dabei muþ ber¸cksichtigt werden, daþ von jeder synsystematischen Ordnung unterschiedlich viele Fl‰chen untersucht wurden.
Lycaena helle, Boloria eunomia
Bei diesen Arten wurden die Daten von 1999 f¸r die Korrelationen verwendet, da in diesem Jahr die Begehungen h‰ufiger und in kleineren Abst‰nden durchgef¸hrt werden konnten.Beiden Faltern ist die ausschlieþliche Raupenfutterpflanze Polygonum bistorta (Schlangen- oder Wiesenknˆterich) gemeinsam. Auþerdem ¸berschneidet sich die Flugzeit ¸ber einen Zeitraum von vier Wochen. Da die Falter auch nebeneinander auf ein und derselben Fl‰che vorkommen, stellt sich die Frage, ob und inwieweit sich ihre jeweiligen autˆkologischen Anspr¸che in Bezug auf die Vegetation unterscheiden. Die kleinr‰umige Bindung an Polygonum bistorta ist bei beiden Arten sehr stark ausgepr‰gt (Tab. 12).
Tab. 12: Bivariable Rangkorrelationen von Lycaena helle und Boloria eunomia mit den Deckungsgraden von Polygonum bistorta nach KENDALL und SPEARMAN (nach SACHS 1997) (Daten von 1999)
Es zeigt sich, daþ die Bindung an den Deckungsgrad von Polygonum bistorta bei Lycaena helle sehr stark ausgepr‰gt ist. Sowohl bei Ber¸cksichtigung aller Fl‰chen als auch bei Ber¸cksichtigung der Fl‰chen mit Vorkommen von Polygonum bistorta ist die Abh‰ngigkeit signifikant. Ðberraschend ist das Ergebnis bei Boloria eunomia. Hier ist die Bindung bei allen Fl‰chen zwar signifikant, doch bei der Korrelation mit den „Polygonum bistorta-Fl‰chen“ ist lediglich eine Tendenz zur Abh‰ngigkeit festzustellen. Das bedeutet, daþ Boloria eunomia zwar abh‰ngig vom Vorkommen nicht aber von der H‰ufigkeit der Futterpflanze ist. Die Aussagen der beiden Korrelationsmethoden stimmen ¸berein. Dadurch werden die Ergebnisse gest¸tzt.
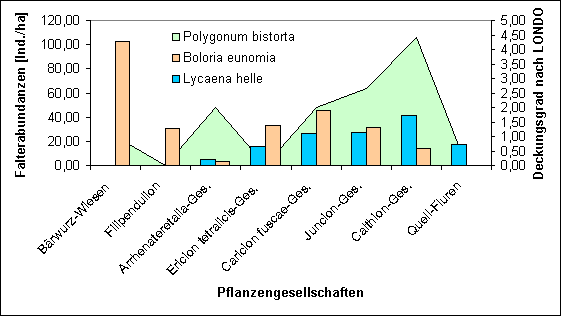
Bei Betrachtung der Verteilung der beiden Arten auf die Vegetationseinheiten ergibt sich ein differenzierteres Bild (Abb.Ý18). Die Verbreitungsschwerpunkte der beiden Arten liegen in unterschiedlichen Bereichen.
So wurde Boloria eunomia ohne Lycaena helle im Filipendulion und auf B‰rwurz-Wiesen gefunden. Auf Quell-Fluren war nur Lycaena helle vertreten, Boloria eunomia fehlte hier. Im Calthion ¸berwiegt Lycaena helle, im Caricion fuscae und Ericion tetralicis ist Boloria eunomia dominanter. Im Juncion und in der Arrhenateretalia-Gesellschaft ist die Abundanz nahezu gleich, wobei in letzterer beide Arten nur sehr schwach vertreten sind.
Lycaena helle – Raupenfunde
Raupen von Lycaena helle (Abb.Ý19) wurden auf zehn Fl‰chen nachgewiesen (L. helleRpos) (Tab. 13). Auf drei dieser Fl‰chen wurden weder im Jahre 1998 noch im Jahre 1999 adulte Falter gefunden. Bei acht weiteren Untersuchungsfl‰chen konnten 1998 zwar adulte Falter nachgewiesen werden, es wurden aber keine Raupen gefunden (L. helleRneg). 1999 verringerte sich die Zahl dieser Fl‰chen auf f¸nf. Tab. 13: Untersuchungsflächen mit Raupen- bzw. Falterfunden von Lycaena helle Es f‰llt auf, daþ der Mittelwert der Falterabundanzen bei den L. helleRpos – Fl‰chen wesentlich hˆher liegt als bei den L. helleRneg – Fl‰chen. Ausgehend von der Annahme, daþ es sich bei den Fl‰chen mit Raupennachweis um „sichere Vorkommen“, d.h. Reproduktionsfl‰chen handelt, ist anzunehmen, daþ hier auch die Abundanz der Falter hˆher liegt. Diese Habitate bieten sowohl den Pr‰imaginalstadien als auch den adulten Faltern die benˆtigten Ressourcen. Eine Ðberpr¸fung der Daten anhand des t-Tests (Tab. 14) ergab jedoch keine Signifikanz dieser Unterschiede (p = 0,45 > 0,05 f¸r 1998, p = 0,31 > 0,05 f¸r 1999). Tab. 14: t-Test für zwei unabhängige Stichproben: Unterschiedlicher Varianz. Ergebnis der Überprüfung zur Abundanzerhöhung in Flächen mit Lycaena helle - Raupen
Boloria selene
Die Raupen von Boloria selene fressen an verschiedenen Veilchen-Arten (Viola canina (Hunds-Veilchen), V. palustris (Sumpf-V.), V. riviniana (Hain-V.), V. hirta (Rauhhaar-V.). Auf den Untersuchungsfl‰chen wurden davon die Arten Viola palustris und Viola canina angetroffen. Die Bindung an die Raupenfutterpflanze Viola palustris ist unter Ber¸cksichtigung aller Fl‰chen signifikant (Tab. 15). Tab. 15: Bivariable Rangkorrelationen von Boloria selene mit den Deckungsgraden von Viola palustris und Viola canina nach KENDALL und SPEARMAN (nach SACHS 1997) (Daten von 1998)
Betrachtet man lediglich die Fl‰chen mit Viola palustris, so ist die Bindung an den Deckungsgrad nicht signifikant. Ebenso verh‰lt es sich bei der Korrelation mit beiden Veilchen-Arten. Bei Viola canina ist die Bindung nicht nachzuweisen. Die Korrelation mit den Deckungsgraden der Pflanze unter Ber¸cksichtigung aller Fl‰chen ergab keine Signifikanz. Da die Art nur auf zwei Fl‰chen nachgewiesen werden konnte, wurde auf eine Korrelation nur mit diesen Arealen verzichtet. Die beiden Veilchenarten kamen auf keiner der Fl‰chen nebeneinander vor. Es kˆnnen also keine Abh‰ngigkeiten bei den verschiedenen Korrelationen bestanden haben. Viola palustris erscheint als die wichtigste Futterpflanze f¸r Boloria selene auf den Untersuchungsfl‰chen. Dabei ist weniger der Deckungsgrad als das Vorkommen der Pflanze ausschlaggebend.
Brenthis inoAuch f¸r Brenthis ino wurden Rangkorrelationen zur Ðberpr¸fung der Abh‰ngigkeit des Vorkommens der Adulten von den Deckungsgraden der Raupenfutterpflanzen durchgef¸hrt (Tab. 16). Wie die Veilchenarten kamen auch Filipendula ulmaria (M‰des¸þ) und Sanguisorba officinalis (Groþer Wiesenknopf) auf keinem der untersuchten Raster gemeinsam vor. Tab. 16: Bivariable Rangkorrelationen von Brenthis ino mit den Deckungsgraden von Filipendula ulmaria und Sanguisorba officinalis nach KENDALL und SPEARMAN (nach SACHS 1997) (Daten von 1998)
F¸r Filipendula ulmaria ergibt sich bei Ber¸cksichtigung aller Fl‰chen eine enge Bindung der Falter. Bei den Fl‰chen mit Filipendula ulmaria ist nur mehr eine Tendenz zur Abh‰ngigkeit zu erkennen. Da Filipendula ulmaria oft fl‰chendeckend vorkommt ist davon auszugehen, das, ‰hnlich zu Boloria eunomia und Polygonum bistorta, ab einem gewissen Deckungsgrad die Falterabundanz nicht mehr zunimmt. Bei Sanguisorba officinalis ist es umgekehrt, die Bindung ist hier nur unter Ber¸cksichtigung der Fl‰chen mit der Pflanzenart zu erkennen. Das mag an den hohen Deckungsgraden von Filipendula ulmaria und den hohen Abundanzen des Falters in diesen Bereichen liegen. Diese wirken sich nat¸rlich auf die Korrelation aller Fl‰chen aus. Bei Betrachtung der Fl‰chen mit beiden Pflanzenarten ist die Korrelation in beiden F‰llen signifikant. Es zeigt sich also, daþ die Falterart auf den untersuchten Fl‰chen von beiden Pflanzen abh‰ngig ist. Die Vermutung, daþ auch Sanguisorba officinalis als Futterpflanze dient, wird verst‰rkt.
Boloria aquilonarisF¸r Boloria aquilonaris konnte aufgrund der geringen Anzahl an Nachweisen (sechs Individuen in vier Rastern) keine Rangkorrelation durchgef¸hrt werden. Drei dieser Raster gehˆren zu einer Untersuchungsfl‰che. In einem Raster wurde die Raupenfutterpflanze Oxycoccus palustris angetroffen. Das vierte Raster gehˆrt zu einer anderen Untersuchungsfl‰che, Oxycoccus palustris wurde zwar nicht auf dem Raster direkt, aber in anderen Bereichen des Areals gefunden. Nach Auskunft von Doerpinghaus handelt es sich bei beiden Fl‰chen um langj‰hrige Habitate des Falters, also wahrscheinlich um Reproduktionsfl‰chen [2] . Drei der Raster sind eindeutig dem Ericion tetralices zuzuordnen, eines f‰llt in den Bereich des Juncion. Auf allen Fl‰chen sind Arten der Heidemoore anzutreffen, vor allem Torfmoose (Sphagnum spec.).
[1] Wagener, F. (1998): M¸ndliche Aukunft. Biologische Station des Kreis Euskirchen Nettersheim. [2] Doerpinghaus, A. (1999): M¸ndliche Auskunft.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||