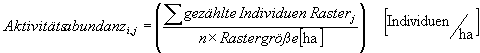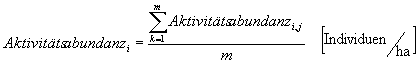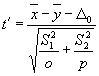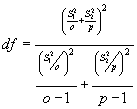Methoden::Erfassung
der Falterfauna
Systematik der „Tagaktiven Schmetterlinge“„Tagaktive Schmetterlinge“ ist kein systematischer Begriff. Im allgemeinen werden hierzu neben den Tagfaltern (Rhopalocera) auch die Widderchen (Zygaenidae) gezählt. Dabei handelt es sich um Gruppen ganz unterschiedlicher systematischer Zuordnung. Die nachfolgende Einordnung in das System folgt Scoble (1992): Die Lepidoptera werden in die Höheren Ditrysia und die Niederen Ditrysia unterteilt. Zur ersten Gruppe zählen die „Großschmetterlinge“ (Makrolepidoptera), deren phylogenetische Beziehungen als unsicher gelten. Das typische Merkmal dieser Gruppe sind die an und auf Pflanzenmaterial fressenden Raupen mit ihren dazu angepaßten Beinpaaren und nicht etwa Größenunterschiede der Falter, wie der Name vortäuscht. Einige Arten haben sich sekundär zu anderen Ernährungstypen wie Bohrer, Parasiten und Fungivoren entwickelt. Zu den Makrolepidoptera zählen die Rhopalocera. Diese unterteilen sich in drei Unterordnungen, von denen zwei, die Papillionoidea (Echte Tagfalter) und die Hesperoidea (Unechte Tagfalter oder Dickkopffalter), in Europa vorkommen. Von der Gruppe der Niederen Ditrysia, mit überwiegend endogener Lebensweise der Raupen, zählt man die Zygaenidae (Widderchen) zu den Tagaktiven Schmetterlingen. Sie zeichnen sich durch extern lebende Raupen und durchweg tagaktive Imagines aus (Prinz & Naumann 1988). Für die Bestimmung der Arten wurden die Werke von Tolman & Lewington (1998) und Weidemann (1995) verwendet. Die Nomenklatur folgt Karsholt & Razowski (1996). Nach diesem Werk richtet sich auch die aktuelle ROTE LISTE (Pretscher et al. 1998).
Erfassung der FalterfaunaDie ausgewählten Flächen wurden ab Anfang Mai regelmäßig begangen, um den Beginn der Imaginalphase von Lycaena helle feststellen zu können.
Linien-TransektmethodeDie Linien-Transektmethode wurde in der Ornithologie von Balogh (1958) begründet und für die Erfassung von Tagfaltern übernommen (Moore 1975, Pollard 1977). Nach der Standardisierung durch Steffny (1982) wurde sie in der Vergangenheit schon mehrmals in der Eifel angewendet (Weidner 1992, Köhler 1993 u. a.). Um eine Vergleichbarkeit mit anderen Untersuchungen zu gewährleisten, müssen folgende Standardbedingungen erfüllt sein:
Diese limitierenden Voraussetzungen schränken die Zahl der Begehungstage ein:
Durch das Verfahren wurden alle adulten tagaktiven Schmetterlinge erfaßt. Hierzu wurden die Untersuchungsflächen in Abständen von ein bis drei Wochen entlang vorher festgelegter Strecken in gleichmäßigem Tempo abgeschritten. Innerhalb einer Entfernung von jeweils 2,5 m links und rechts dieser Strecke wurden gesichtete Tagfalter bestimmt (teilweise unter Hilfe eines Monokulars: Zeiss 5 x 10). Aufgeschreckte Individuen und schwer bestimmbare Arten wurden mit einem Schmetterlingsnetz (Ø 40 cm, 70 cm-Netz) möglichst schonend gefangen, „in der Hand“ bestimmt und freigelassen. Es ergeben sich Transekte von 5 m Breite und einer von der Größe der Fläche abhängigen Länge. Die Auswahl der Strecken richtet sich primär nach den zu erwartenden Faltern und damit nach der Vegetation und der Beschattung der Untersuchungsfläche. Ein weiteres Kriterium ist die Form des Transekts: es sollte möglichst gerade sein und keine rückläufigen Abschnitte aufweisen, um Doppelzählungen zu vermeiden. Die gesichteten Falter und ihre Aktivität (Flug, Ruhe, Nektaraufnahme mit Nektarpflanze, Kopulation, Eiablage) wurden auf einem Erfassungsbogen vermerkt.
Vor jeder Begehung wurden zusätzlich folgende Daten notiert:
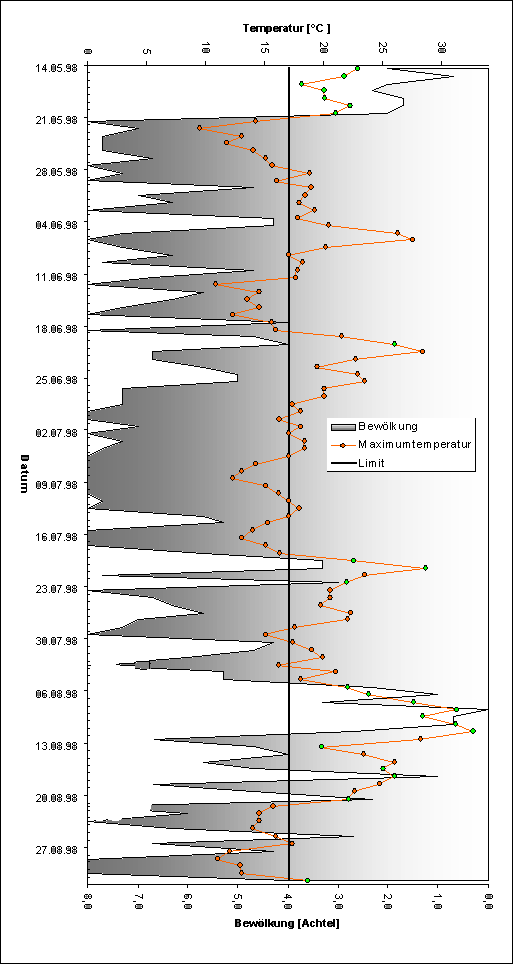
Die Messung der Temperatur erfolgte in °C mit einem handelsüblichen Digitalthermometer, die Windstärke wurde anhand der Beaufort-Skala ermittelt und der Bewölkungsgrad in Achteln geschätzt. Um eine bessere Auflösung der Erfassung zu erreichen, wurden die Transekte in bis zu 3 Raster unterteilt. Diese Raster unterschieden sich in ihrer Vegetation; alle anderen Parameter, wie Exposition, Neigung, angrenzende Pflanzengesellschaften usw. waren nahezu gleich. Unterschiede in der Artzusammensetzung der Falterfauna eines Transekts sollten so in erster Linie auf die unterschiedlichen Vegetationseinheiten zurückzuführen sein. Durch die Einteilung in Mikroraster wurde die Vergleichbarkeit der Ergebnisse mit denen anderer Untersuchungen nicht beeinträchtigt. Es ist möglich, die Daten sowohl nach den Rastern als auch nach den Linientransekten auszuwerten. Im Jahre 1998 wurden die Flächen von Mitte Mai (Beginn der Flugzeit von Lycaena helle) bis August (Ende der Flugzeit von Boloria aquilonaris) begangen. 1999 richtete sich die Untersuchung lediglich nach den Flugzeiten von Lycaena helle und Boloria eunomia und endete im Juli.
Erfassung von PräimaginalstadienDer Nachweis von adulten Faltern auf einer Fläche bedeutet nicht, daß es sich hier um ein dauerhaftes Habitat dieser Art handeln muß. Es besteht immer die Möglichkeit der aktiven Zuwanderung oder der Windverdriftung. Bei den Präimaginalstadien gibt es diese Möglichkeiten nicht, ihr Nachweis deutet in wesentlich höherem Maße auf ein gesichertes Vorkommen bzw. auf eine geeignete Fortpflanzungs- und Entwicklungsstätte hin. Das Raupen- und insbesondere das immobile Puppenstadium sind die kritischsten Phasen im Lebenszyklus der Lepidoptera. Während die Adulten pessimalen Umweltbedingungen wie Nahrungsmangel, Räuber- und Parasitendruck oder extremen Feuchte- und Temperaturverhältnissen durch Ortswechsel begegnen können, müssen die Präimaginalstadien diese Einflüsse an ihrem Standort ertragen. „Qualität und Ausdehnung geeigneter Entwicklungsstätten sind für Vorkommen und Häufigkeit der meisten Tagfalter weit wichtiger als die von Laien vielfach überbewerteten Saughabitate“ (Hermann 1998). Da die Abundanz der vorhandenen Präimaginalstadien einer Art in der Regel um das 10- bis 100fache höher ist als die der Imagines, liegt es nahe, sich diesen Umstand bei Bestandsaufnahmen zunutze zu machen. Aufgrund von Unterschieden bei der Eiablage, dem Raupenverhalten und dem Verpuppungsort eignen sich jedoch nicht alle Arten gleichermaßen zur Erfassung von Präimaginalstadien (Tab. 1). Aus diesem Grunde wurde lediglich gezielt nach den Raupen von Lycaena helle gesucht. Tab. 1 : Effizienz und Notwendigkeit der gezielten Suche von Präimaginalstadien von Boloria aquilonaris, Boloria eunomia, Boloria selene, Brenthis ino und Lycaena helle (nach Hermann 1998)
Der sicherste Nachweis einer Entwicklungsstätte wäre die Erfassung von gerade schlüpfenden Tieren aus der Puppenhülle. Dadurch würde der gesamte Entwicklungszyklus von einer Generation zur nächsten auf der Beobachtungsfläche bewiesen. Das ist in der Praxis jedoch nicht möglich. Eine Näherung wäre die Erfassung von Puppen. Von den in Tab. 1 genannten Arten sind die Puppen aber nur sehr schwer zu finden. Daher beschränkte sich die Suche nach Präimaginalstadien auf kurz vor der Verpuppung stehende Larven von Lycaena helle (Ende Juli bis Mitte August 1998). Die Raupenfutterpflanze von Lycaena helle ist Polygonum bistorta. Zum Nachweis wurden Bestände dieser Pflanze gezielt nach den Raupen durchsucht. Die Auswahl der Bereiche richtete sich nach den Arbeiten von Drews & Fechner (1996) und Bück (1996), in denen Eiablageplätze und Entwicklungsstätten von Lycaena helle charakterisiert werden. Pro Untersuchungsfläche wurden drei Stunden Suchzeit veranschlagt. Bei Fund von mindestens zwei Raupen wurde die Fläche als Raupe-positiv (L. helleRpos) eingestuft und die Suche abgebrochen. Wenn nach drei Stunden keine Raupe nachgewiesen werden konnte, wurde die Fläche als Raupe-negativ (L. helleRneg) eingestuft und die Suche beendet.
Ermittlung der potentiellen SonnenscheindauerDie vom Deutschen Wetterdienst (1999) übernommenen Daten reichen zur Charakterisierung der örtlichen Situation bezüglich der Sonnenscheindauer nicht aus. Je nach Beschaffenheit der Umgebung einer Untersuchungsfläche kommt es zu mehr oder weniger starker Beschattung. Ein auf einer Hochebene gelegenes Heidemoor wird naturgemäß besser von der Sonnenstrahlung erreicht, als ein eingekerbtes Bachtal, in dem die Beschattung bei niedrigem Sonnenstand in den Wintermonaten sogar 100 % betragen kann. Dadurch kommt es zu großen Differenzen im Mesoklima der untersuchten Bereiche. Das wirkt sich indirekt (über Unterschiede in der Vegetation) und direkt auf die Fauna und damit auf alle Entwicklungsstadien der Falter aus. Um diese Unterschiede zu quantifizieren, wurde von jeder Probefläche die potentielle Sonnenscheindauer der Monate Juni und Juli ermittelt. Mit dem Horizontoskop nach Tonne, welches auf einem 40 cm hohen Holzstativ aufgestellt, mit einer Wasserwaage horizontal eingemessen und mit einem Kompaß eingenordet wird, kann die Besonnung zu einer bestimmten Jahreszeit abgelesen werden. Der Horizont mit eventuell beschattenden Strukturen wird bei diesem Gerät auf ein durchsichtiges Kunststoffgewölbe projiziert. Innerhalb des Gewölbes kann auf einer je nach geographischer Breite austauschbaren Schablone der Sonnenlauf einer Jahreszeit abgelesen werden. Die potentielle Sonnenscheindauer ergibt sich dann durch die Schnittpunkte des projizierten Horizonts und der Linie des Sonnenlaufs auf der Schablone. Die Messung erfolgte in repräsentativen Bereichen (meist in der Mitte) eines jeden Transekts. Bei der Auswahl des Meßortes wurde darauf geachtet, daß für die Bestrahlung der Gesamtfläche unwesentliche Strukturen, wie einzelne Bäume oder Gebüsche, keinen Einfluß auf die Messung hatten. Der ermittelte Wert sagt nichts über die tatsächliche direkte Sonneneinstrahlung aus. Berücksichtigt man aber, daß innerhalb des recht kleinräumigen Untersuchungsgebiets die Bewölkung auf allen Probeflächen während des Tageslaufs nahezu gleich ist, so kann man die potentielle Sonnenscheindauer als relative Größe zum direkten Vergleich annehmen. Es ist nicht zu entscheiden, ob eine ermittelte Verschiedenartigkeit der Falterfauna zweier Flächen direkt auf Unterschiede bezüglich der Sonneneinstrahlung oder auf die dadurch hervorgerufene Verschiedenheit der Vegetation zurückzuführen ist. Eine direkte Gegenüberstellung der Ergebnisse der Falterkartierung und der Messung der potentiellen Sonnenscheindauer muß daher immer im Zusammenhang mit der vorhandenen Vegetation betrachtet werden.
Berechnungsmethoden für verschiedene Indices und statistische VerfahrenDie folgenden Formeln werden benötigt, um aus den Rohdaten der einzelnen Flächen vergleichbare Werte zu ermitteln. Die Abundanz ist die Anzahl der Individuen pro Flächeneinheit. Die Verteilung der Individuen auf die Arten wird durch den Dominanzindex angegeben. Die Präsenz gibt an, in welchem Anteil der Untersuchungsflächen eine Art vorkommt.
Formel 1 : Aktivitätsabundanz der Arti pro ha im Rasterj
Formel 2 : Aktivitätsabundanz der Arti pro ha in der Summe der Untersuchungsflächen
Formel 3 : Dominanzindex d der Arti in der Summe der Untersuchungsflächen
Tab. 2: Dominanzklassen nach ENGELMANN (1978)
Formel 4 : Präsenz der Art i im Untersuchungsgebiet n = Anzahl der Begehungen m = Anzahl der Raster
Der Diversitätsindex nach Shannon-Weaver beschreibt den mittleren Grad der Ungewißheit, irgendeine bestimmte Art bei zufälliger Probeentnahme anzutreffen.
Die Evenness dient als Vergleichsmaß für die Diversitätsindizes verschiedener Ökosysteme. HS = Diversität bezogen auf Artenzahlen S = Gesamtzahl der Arten pi = Wahrscheinlichkeit des Auftretens der Art i N = Gesamtindividuenzahl ni = Individuenzahl der Art i
Der t-Test dient zur Überprüfung von zwei Datenreihen auf zufällige oder nichtzufällige Unterschiede des Mittelwertes. Dabei ist:
o = Stichprobenzahl der ersten Datenreihe; p = Stichprobenzahl der zweiten Datenreihe; S1 = Standardabweichung der ersten Datenreihe; S2 = Standardabweichung der zweiten Datenreihe; D0 = Differenz der Standardabweichungen.
Formel 7 : Zweistichproben t-Test: Unterschiedliche Varianzen
Formel 8 : Näherungswert für den Freiheitsgrad des Zweistichproben t-Test: Unterschiedliche Varianzen
Erfassung der VegetationVegetationsaufnahmenDie Nomenklatur der Pflanzen richtet sich nach Rothmaler (1994). Die Aufnahme der Vegetation in den Rastern der Transekte wurde von A. Doerpinghaus nach der Methode von Braun-Blanquet (1964) in den Jahren 1997 und 1998 durchgeführt. Die Deckungsgrade der Pflanzenarten wurden in Einheiten der Londo-Skala (nach Dierschke 1994) übernommen.
Auswertung der AufnahmedatenDie Raster der Untersuchungsflächen wurden anhand der Vegetationsaufnahmen nach Oberdorfer (1994) pflanzensoziologisch zugeordnet. Eine Zuordnung bis zur Ebene der Assoziation war in vielen Fällen aufgrund der Gesellschaftsübergänge und anthropogener Einflüsse nicht möglich. Hier wurde die Zugehörigkeit zum Verband oder der höheren sytaxonomischen Einheit angegeben. EDVDie Auswertung der Daten, einschließlich der deskriptiven Statistik und statistischer Tests wurde mit den Programm Excel 97 der Firma Microsoft durchgeführt. Mit dem Programm SPSS 6.1 wurden die Rangkorrelationen berechnet. Die Bildbearbeitung erfolgte mit dem Programm Photoshop 5.0 der Firma Adobe. |
||||||||||||||||||||||||||||||||